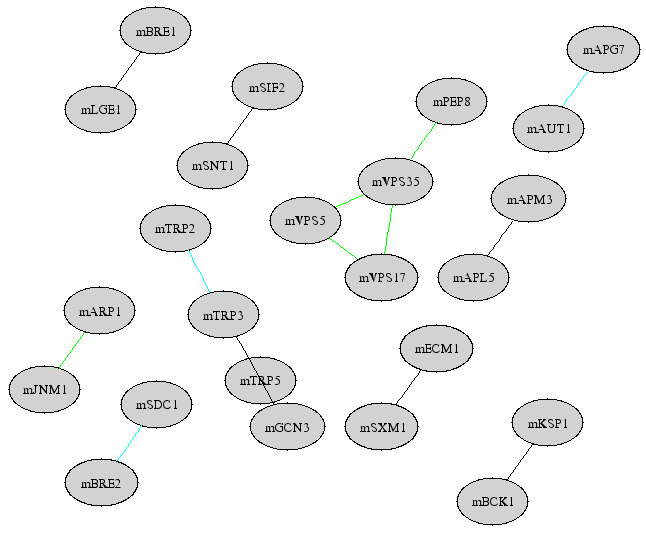
Module number 3267
Database revision : gnsdb28.10
Date : Tue Feb 25 17:22:29 2003
How to read this figure?
this is an automaticly generated SAMBA report
Computational Genomics Lab, Tel-Aviv uniresity